

|
The emergence of chordates are the first step that nature made to put rigidity in lifeforms. Having a structure seems to be advantageous along the evolutionary path as this was later joined by the development of exoskeletons. by invertebrates It also allow for creatures to emerge beyond the buoyant world of the oceans...
...and take our first steps onto solid ground. |
"There's safety only in a stiff backbone."
Water Margin by Shi Nai'an, Chapter 24
The Phylum Chordata includes the well-known vertebrates (fishes, amphibians, reptiles, birds, mammals). The vertebrates and hagfishes together comprise the taxon Craniata. The remaining chordates are the tunicates (Urochordata), lancelets (Cephalochordata), and, possibly, some odd extinct groups. With few exceptions, chordates are active animals with bilaterally symmetric bodies that are longitudinally differentiated into head, trunk and tail. The most distinctive morphological features of chordates are the notochord, nerve cord, and visceral clefts and arches.

Lancelets, a Cephalochordata
|

Sea squirts, a Tunicata
|

Blue whale, a Craniate
|
Chordates are well represented in marine, freshwater and terrestrial habitats from the Equator to the high northern and southern latitudes. The oldest fossil chordates are of Cambrian age. The earliest is Yunnanozoon lividum from the Early Cambrian, 525 Ma (= million years ago), of China. This was just recently described and placed with the cephalochordates (Chen et al., 1995). Another possible cephalochordate is Pikaia (Nelson, 1994) from the Middle Cambrian. These fossils are highly significant because they imply the contemporary existence of the tunicates and craniates in the Early Cambrian during the so-called Cambrian Explosion of animal life. Two other extinct Cambrian taxa, the calcichordates and conodonts, are uncertainly related to other Chordata (Nelson, 1994). In the Tree of Life project, conodonts are placed as a subgroup of vertebrates.
Chordates other than craniates include entirely aquatic forms. The strictly marine Urochordata or Tunicata are commonly known as tunicates, sea squirts, and salps. There are roughly 1,600 species of urochordates; most are small solitary animals but some are colonial, organisms. Nearly all are sessile as adults but they have free-swimming, active larval forms. Urochordates are unknown as fossils. Cephalochordata are also known as amphioxus and lancelets. The group contains only about 20 species of sand-burrowing marine creatures. The Cambrian fossils Yunnanozoon and Pikaia are likely related to modern cephalochordates.
During the Ordovician Period (510 - 439 Ma) jawless or agnathan fishes appeared and diversified. These are the earliest known members of Vertebrata, the chordate subgroup that is most familiar to us. Fossils representing most major lineages of fish-like vertebrates and the earliest tetrapods (Amphibia) were in existence before the end of the Devonian Period (363 Ma). Reptile-like tetrapods originated during the Carboniferous (363 - 290 Ma), mammals differentiated before the end of the Triassic (208 Ma) and birds before the end of the Jurassic (146 Ma).
The smallest chordates (e.g. some of the tunicates and gobioid fishes) are mature at a length of about 1 cm, whereas the largest animals that have ever existed are chordates: some sauropod dinosaurs reached more than 20 m and living blue whales grow to about 30m.
Chordates other than craniates include entirely aquatic forms. The strictly marine Urochordata or Tunicata are commonly known as tunicates, sea squirts, and salps. There are roughly 1,600 species of urochordates; most are small solitary animals but some are colonial, organisms. Nearly all are sessile as adults but they have free-swimming, active larval forms. Urochordates are unknown as fossils. Cephalochordata are also known as amphioxus and lancelets. The group contains only about 20 species of sand-burrowing marine creatures. The Cambrian fossils Yunnanozoon and Pikaia are likely related to modern cephalochordates.
During the Ordovician Period (510 - 439 Ma) jawless or agnathan fishes appeared and diversified. These are the earliest known members of Vertebrata, the chordate subgroup that is most familiar to us. Fossils representing most major lineages of fish-like vertebrates and the earliest tetrapods (Amphibia) were in existence before the end of the Devonian Period (363 Ma). Reptile-like tetrapods originated during the Carboniferous (363 - 290 Ma), mammals differentiated before the end of the Triassic (208 Ma) and birds before the end of the Jurassic (146 Ma).
The smallest chordates (e.g. some of the tunicates and gobioid fishes) are mature at a length of about 1 cm, whereas the largest animals that have ever existed are chordates: some sauropod dinosaurs reached more than 20 m and living blue whales grow to about 30m.
|
Shared Characteristics
The notochord is an elongate, rod-like, skeletal structure dorsal to the gut tube and ventral to the nerve cord. The notochord should not be confused with the backbone or vertebral column of most adult vertebrates. The notochord appears early in embryogeny and plays an important role in promoting or organizing the embryonic development of nearby structures. In most adult chordates the notochord disappears or becomes highly modified. In some non-vertebrate chordates and fishes the notochord persists as a laterally flexible but incompressible skeletal rod that prevents telescopic collapse of the body during swimming. The nerve cord of chordates develops dorsally in the body as a hollow tube above the notochord. In most species it differentiates in embryogeny into the brain anteriorly and spinal cord that runs through the trunk and tail. Together the brain and spinal cord are the central nervous system to which peripheral sensory and motor nerves connect. The visceral (also called pharyngeal or gill) clefts and arches are located in the pharyngeal part of the digestive tract behind the oral cavity and anterior to the esophagus. The visceral clefts appear as several pairs of pouches that push outward from the lateral walls of the pharynx eventually to reach the surface to form the clefts. Thus the clefts are continuous, slit-like passages connecting the pharynx to the exterior. The soft and skeletal tissues between adjacent clefts are the visceral arches. The embryonic fate of the clefts and slits varies greatly depending on the taxonomic subgroup. In many of the non-vertebrate chordates, such as tunicates and cephalochordates, the clefts and arches are elaborated as straining devices concerned with capture of small food particles from water. |
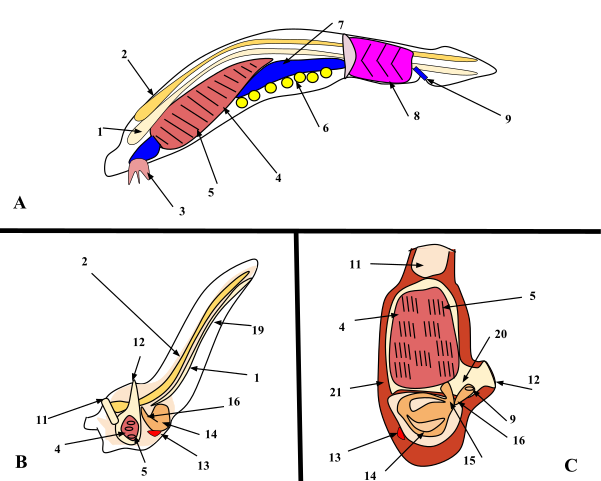
1. Notochord, 2. Nerve chord, 3. Buccal cirri, 4. Pharynx, 5. Gill slit, 6. Gonad, 7. Gut, 8. V-shaped muscles, 9. Anus, 10. Inhalant syphon, 11. Exhalant syphon, 12. Heart, 13. Stomach, 14. Esophagus, 15. Intestines, 16. Tail, 17. Atrium, 18. Tunic
|
In typical fish-like vertebrates and juvenile amphibians the walls of the pharyngeal clefts develop into gills that are organs of gas exchange between the water and blood. In adult amphibians and the amniote tetrapods (= reptiles, birds and mammals) the anteriormost cleft transforms into the auditory (Eustachian) tube and middle ear chamber, whereas the other clefts disappear after making some important contributions to glands and lymphatic tissues in the throat region. The skeleton and muscles of the visceral arches are the source of a great diversity of adult structures in the vertebrates. For example, in humans (and other mammals) visceral arch derivatives include the jaw and facial muscles, the embryonic cartilaginous skeleton of the lower jaw, the alisphenoid bone in the side wall of the braincase, the three middle ear ossicles (malleus, incus and stapes), the skeleton and some musculature of the tongue, the skeleton and muscles of the larynx, and the cartilaginous tracheal rings.
Evolution of Chordates
The fossil record of chordates begins in the early Cambrian period, roughly 530 million years ago. The oldest known chordate fossil was found in China and described in 1995. It is a member of the species Yunnanozoon lividium, within the subphylum Cephalochordata. The earliest fossil evidence for members of the vertebrate subphylum also dates back roughly 530 million years. This is when fossils of the jawless fish first appeared. The earliest tetrapods (having four legs) are the amphibians, with fossils dating back to the Devonian period, roughly 363 million years ago. Shortly thereafter, reptile-like tetrapods arose. It was some time after the appearance of reptiles, approximately 80 million years, that mammals entered the scene. The first mammalian fossils date back to the Triassic period, about 208 million years ago. Finally, bird fossils appear 146 million years ago in the Jurassic period.
Chordate evolution, like that of arthropods, includes major habitat transitions. The earliest chordates were all marine animals like tunicates and lancelets. As chordates continued to evolve, they spread to freshwater habitats and ultimately to land. The amphibians represent an intermediate phase in the water to land transition of chordates. The evolution of birds further increased the distribution of chordates by expanding their populations into aerial habitats.
The evolution of chordates led to a variety of specialized structural features and behavioral modifications that were necessitated by the environmental pressures imposed on different groups within the phylum. Vertebrates in particular show an enormous range of features that represent different evolutionary modifications of the same original structure. One example of this is the bird beak.
Many evolutionary adaptations in vertebrates are related to their increased size, relative to earlier chordates, and an increased level of activity. For example, a strong backbone, an extensive endoskeleton, and a protective covering for the brain (skull) are necessary to provide structural support and internal protection to a large fast-moving animal. In addition, being able to move rapidly while navigating a complex environment requires a high level of sensory capacity and a highly developed nervous system to process that sensory information and mediate appropriate responses. The evolution of a large brain and a centralized nervous system accommodated this adaptation. But how did all of this complexity first begin? In order to answer this question we will next examine the earliest chordate species and their proposed origins.
The fossil record of chordates begins in the early Cambrian period, roughly 530 million years ago. The oldest known chordate fossil was found in China and described in 1995. It is a member of the species Yunnanozoon lividium, within the subphylum Cephalochordata. The earliest fossil evidence for members of the vertebrate subphylum also dates back roughly 530 million years. This is when fossils of the jawless fish first appeared. The earliest tetrapods (having four legs) are the amphibians, with fossils dating back to the Devonian period, roughly 363 million years ago. Shortly thereafter, reptile-like tetrapods arose. It was some time after the appearance of reptiles, approximately 80 million years, that mammals entered the scene. The first mammalian fossils date back to the Triassic period, about 208 million years ago. Finally, bird fossils appear 146 million years ago in the Jurassic period.
Chordate evolution, like that of arthropods, includes major habitat transitions. The earliest chordates were all marine animals like tunicates and lancelets. As chordates continued to evolve, they spread to freshwater habitats and ultimately to land. The amphibians represent an intermediate phase in the water to land transition of chordates. The evolution of birds further increased the distribution of chordates by expanding their populations into aerial habitats.
The evolution of chordates led to a variety of specialized structural features and behavioral modifications that were necessitated by the environmental pressures imposed on different groups within the phylum. Vertebrates in particular show an enormous range of features that represent different evolutionary modifications of the same original structure. One example of this is the bird beak.
Many evolutionary adaptations in vertebrates are related to their increased size, relative to earlier chordates, and an increased level of activity. For example, a strong backbone, an extensive endoskeleton, and a protective covering for the brain (skull) are necessary to provide structural support and internal protection to a large fast-moving animal. In addition, being able to move rapidly while navigating a complex environment requires a high level of sensory capacity and a highly developed nervous system to process that sensory information and mediate appropriate responses. The evolution of a large brain and a centralized nervous system accommodated this adaptation. But how did all of this complexity first begin? In order to answer this question we will next examine the earliest chordate species and their proposed origins.
Ponder this
Rigidity is not required in an aquatic environment, and is often disadvantageous. Why didn't vertebrates simply die out immediately after developing these characteristic?
Why do some invertebrates (insects, mollusks, crustaceans) develop exoskeletons rather than vertebrae?
Discuss
What advantages do vertebrates have over invertebrates? How about the other way around? At the period and place where the earliest vertebrates appear, what conditions allow for them to develop these evolutionary advantages? Why don't the invertebrates of that period and place go extinct?
Further reading
Chordate, from the Encyclopedia of Life
"Chordate evolution and the three-phylum system", from the Proceedings of the Royal Society
"Biology of Invertebrate Chordates", from ThoughtCo








